






國自然熱點-遷移體的最近新文章:PI(4,5)P -Rab35軸調節遷移體形成

遷移體是最近發現的新興細胞器,其形成與細胞遷移密切相關。當細胞在胞外基質上遷移時,被稱為收縮纖維(RFs)的長膜突起從細胞后部的質膜上拉出。RFs就像連接細胞體和基質的繩索。遷移體是一種大的囊泡狀結構,在RFS形成開始數小時后開始在RFs上生長。遷移體內部包含多個小囊泡,像石榴一樣,可攜帶多種生物分子。遷移體提供了一種整合和傳遞時空化學信息的機制,用于細胞間的交流。目前關于遷移體的中標項目逐漸增多,遷移體的功能和機理研究仍待發掘,因此不失為基金申請的好方向。下圖為2023年部分遷移體中標項目的題目:
那么,如何在課題中引入和研究遷移體呢?下面,作者通過一篇最近的新高分文章解讀來了解。
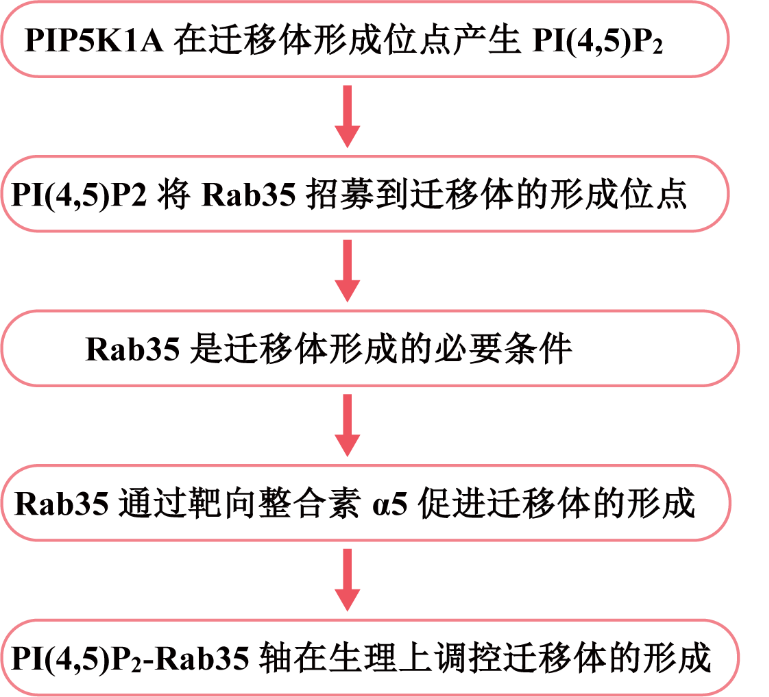
遷移體是最近發現的細胞器,它形成于遷移細胞后緣的RFs的末端或分支點上。先前,作者發現整合素在遷移體形成部位的募集對遷移體的生物發生至關重要。在這項研究中,作者發現在遷移體形成之前,PIP5K1A(一種將PI4P轉化為PI(4,5)P2的PI4P激酶)被募集到遷移體形成位點。PIP5K1A的募集導致在遷移體形成位點產生PI(4,5)P2。一旦積累,PI(4,5)P2通過與Rab35的C端多堿性簇相互作用將Rab35招募到遷移體形成位點。作者進一步證明,活性Rab35通過在遷移體形成位點募集和聚集整合素α5來促進遷移體的形成,這可能是由整合素α5和Rab35之間的相互作用介導。該研究確定了協調遷移體生物形成的上游信號通路,于2023年8月發表在《Journal of Cell Biology》,IF = 44.1。
研究思路
主要研究結果
1. PIP5K1A在遷移體形成位點產生PI(4,5)P2
之前,作者發現PI(4,5)P2的探針PLCγ-PH-TagBFP可以標記遷移體,作者在這里證實了這一點(圖1A)。使用抗PI(4,5)P2抗體對細胞進行染色也顯示遷移體中PI(4,5)P2信號的富集(圖1B)。這些結果表明遷移體含有PI(4,5)P2。為了研究PI(4,5)P2在遷移體上的動態分布,作者進行了延時成像發現PI(4,5)P2的探針PLCγ-PH-TagBFP在TSPAN4之前被募集到遷移體(圖1C)。接下來,作者比較了PLCγ-PH-TagBFP與整合素α5的動態關系。作者發現PLCγ-PH-TagBFP的募集速度略快于整合素α5(圖1D, E)。這些數據表明在遷移體生長之前,PI(4,5)P2在遷移體形成部位產生或聚集。PI4P激酶可將PI4P轉化為PI(4,5)P2,從而產生PI(4,5)P2。為了檢測PI4P激酶是否參與遷移體形成部位PI(4,5)P2的生成,作者對細胞進行了染色,作者發現PIP5K1A定位在遷移體上(圖1F)。同樣,異位表達的PIP5K1A-GFP定位在遷移體上,延時成像顯示PIP5K1A-GFP被招募到遷移體形成的位置,比TSPAN4的招募要早(圖1G)。這與PI(4,5)P2信號的外觀一致(圖1C)。
接下來,作者測試了PIP5K1A是否在遷移體形成位點負責PI(4,5)P2的生成。使用PIP5K1A抑制劑ISA2011B處理細胞,發現ISA2011B處理阻斷了遷移體的形成(圖1H, I)。為了進一步證實PI(4,5)P2是遷移體形成所必需的,在PIP5K1A敲除(KO)細胞中,遷移體的形成明顯減少,而異位表達的野生型(WT) PIP5K1A恢復了遷移體的形成(圖1J, K)。這表明PIP5K1A的酶活性是遷移體形成所必需的,并進一步證明PI(4,5)P2的水平是遷移體形成的決定因素。總之,這些結果表明PIP5K1A在遷移體形成部位產生PI(4,5)P2是遷移體生物發生所必需的。由于PI(4,5)P2可以被脂質磷酸酶水解,作者接下來研究了已知PI(4,5)P2磷酸酶的定位,發現PLCD3定位在遷移體上為了進一步測試PI(4,5)P2在遷移體形成中的作用,作者構建了PLCD3 KO細胞系。在該細胞系中,遷移體的形成明顯增強,PLCD3的異位表達使遷移體的形成恢復到正常水平。總之,這些證實PI(4,5)P2在遷移體形成中的作用。
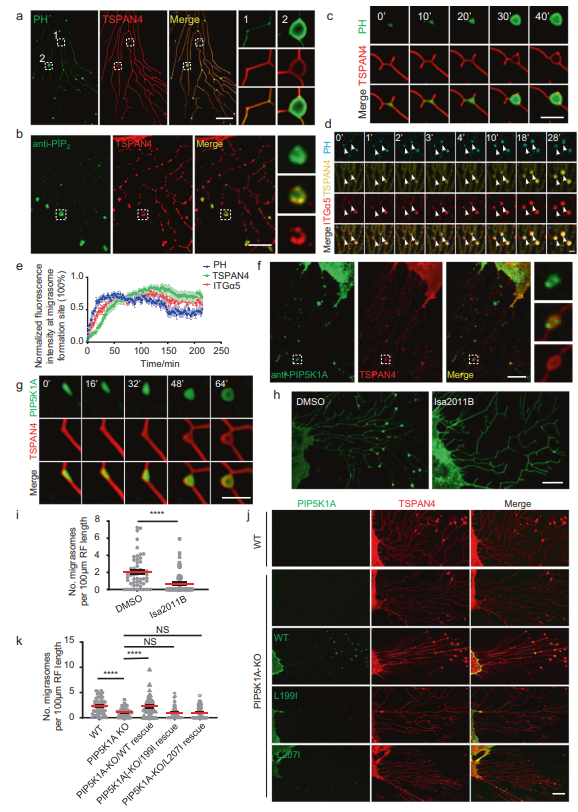
圖1 PIP5K1A在遷移體形成位點產生PI(4,5)P2。
2. PI(4,5)P2將Rab35招募到遷移體的形成位點
接下來,作者研究了PI(4,5)P2如何調控遷移體的形成。作者推斷PI(4,5)P2可能通過募集PI(4,5)P2結合蛋白調控遷移體的形成,而這些蛋白是遷移體形成所必需的。為了篩選可能定位于遷移體的PI(4,5)P2結合蛋白,作者首先編制了大鼠基因組中所有PI(4,5)P2結合蛋白的列表。接下來,作者將此列表與遷移體中的蛋白質列表進行比較,作者之前通過純化遷移體的質譜(MS)分析發現了23個PI(4,5) P2結合蛋白MS列表,包括Rab35(圖2a)。然后,作者為其中的19個蛋白生成了mcherry標記的構建體,并發現其中一些蛋白定位在遷移體上。在這些蛋白中,作者選擇Rab35進行進一步的研究,因為Rab35在細胞器生物發生中起著關鍵作用。
首先用抗Rab35抗體染色細胞來證實Rab35的募集。作者發現內源性Rab35確實定位于遷移體和沿著RFs的小點上(圖2B)。同樣,異位表達的mCherry-Rab35定位于遷移體和遷移體形成位點(圖2C)。為了研究Rab35招募的動態,作者使用mCherry-Rab35進行了延時成像(圖2D),發現Rab35信號首先沿RFs均勻擴散分布。在遷移體形成之前,Rab35信號逐漸集中在分支點,并變得更加強烈。最終,Rab35陽性點開始擴大并長成遷移體(圖2D)。這些結果表明Rab35在遷移體生物發生之前被募集到遷移體形成的位點。接下來,作者測試了Rab35在遷移體形成位點的募集是否依賴于PI(4,5)P2。作者用PIP5K1A抑制劑ISA2011B處理細胞,觀察到Rab35在遷移體形成位點的募集受損(圖2E, F)。同樣,在PIP5K1A KO細胞中,Rab35未能被募集到遷移體形成位點(圖2G, H)。這些結果證實Rab35的募集依賴于PI(4,5)P2。
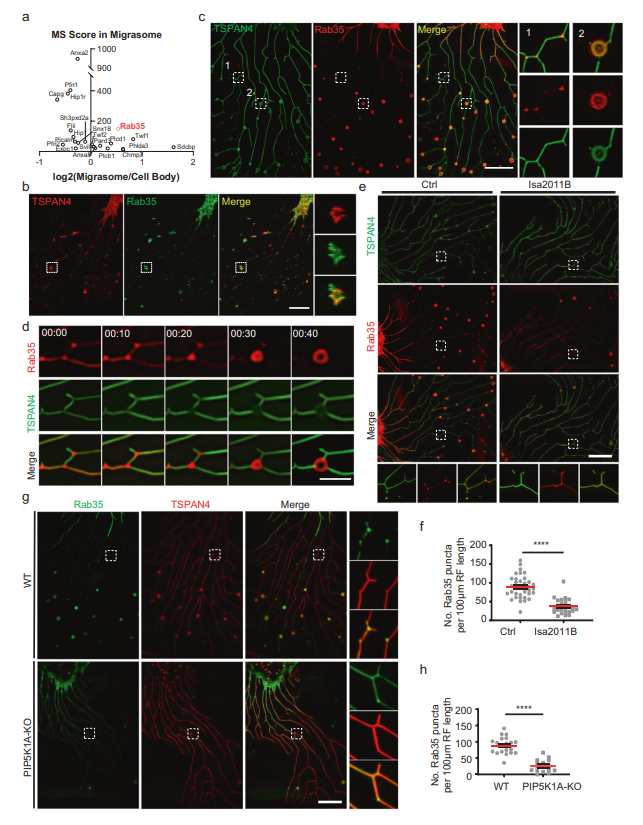
圖2 PI(4,5)P2將Rab35招募到遷移體的形成位點
3. Rab35是遷移體形成的必要條件
接下來,為了測試Rab35是否需要遷移體的形成,作者產生了Rab35 KO細胞系。作者發現,Rab35的KO嚴重損害了遷移體的形成(圖3A, B)。有趣的是,Rab35的KO增加了RFs的數量和長度(圖3A, C)。Rab35 KO細胞系中WT Rab35和組成型活性Rab35-q67l的穩定表達挽救了遷移體的形成,而Rab35-s22n的顯性陰性突變體的穩定表達未能挽救遷移體的形成(圖3D, F)。這些結果表明,活性Rab35是遷移體形成所必需的。先前的文獻表明,PI(4,5)P2通過與其C端多堿性氨基酸團簇(由一段帶正電的賴氨酸和精氨酸殘基組成)相互作用將Rab35招募到質膜上當作者用非極性中性氨基酸Ala (Rab35-7a)取代多堿性氨基酸簇時,作者發現突變體Rab35不能被招募到遷移體中,也不能挽救遷移體的形成(圖3D, E)。為了進一步證實Rab35在遷移體形成中的作用,作者建立了穩定表達WT Rab35、顯性陰性Rab35-s22n和組成活性Rab35-q67l的細胞系。與拯救實驗一致,作者發現過表達WT Rab35和組成活性Rab35-q67l增強了遷移體的形成,而表達顯性陰性的Rab35-s22n減少了遷移體的形成。此外,表達顯性陰性的Rab35-s22n增強遷移體形成(圖3F)。綜上所述,這些數據表明Rab35在調節RFs長度和遷移體形成中起重要作用。
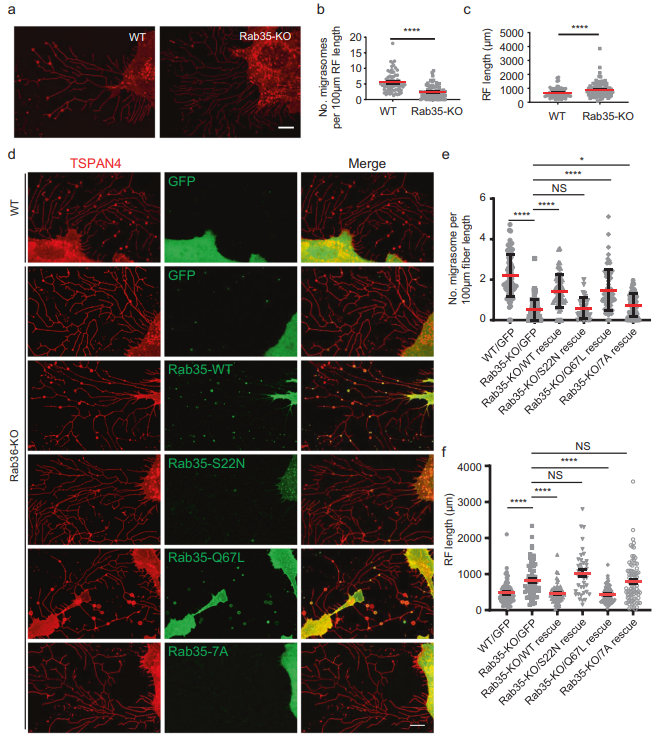
圖3 Rab35是遷移體形成所必需的
4. Rab35通過靶向整合素α5促進遷移體的形成
最后,作者研究了Rab35如何促進遷移體的形成。先前的文獻表明Rab35是整合素運輸所必需的,因此作者選擇研究整合素和Rab35之間的關系。先前作者發現整合素異源二聚體與細胞外基質(ECM)蛋白的配對決定遷移體的形成具體來說,作者發現當細胞在包被特定ECM蛋白的培養皿中生長時,可以與特定ECM蛋白結合的整合素異二聚體在遷移體形成位點高度富集,并在遷移體的生物發生中發揮重要作用。在纖維連接蛋白上生長的細胞中,整合素α5β1在遷移體中高度富集。在WT細胞中,大部分整合素α5集中在遷移體上,縮回纖維中很少有整合素α5。相反,作者發現在Rab35 KO細胞中,ITGα5-GFP不是集中在遷移體形成部位,而是沿RFs均勻彌散分布(圖4A-C)。這表明在Rab35缺失的情況下,整合素對遷移體形成位點的靶向作用受損。接下來,作者研究了rab35依賴性募集ITGα5-GFP的分子機制。先前的報道表明,所有整合素α亞基都可以通過保守的膜-近端GFFKR基序與Rab21結合,該基序也存在于整合素α5.14中。作者想知道Rab35是否可以通過該基序與整合素α5結合。為了驗證這一假設,作者制作了一個整合素α5突變蛋白,GFFKR突變為AAAAA (1-5A)。作為對照,作者還生成了另外4個突變體,其中整合素α5細胞質部分的4組連續5個氨基酸突變為AAAAA (2-5A, 3-5A, 4-5A, 5-5A)(圖4D)。總之,這些突變體覆蓋了大部分整合素α5細胞質結構域。作者發現GFFKR/AAAAA (1-5A)突變體對遷移體形成位點的靶向性受損(圖4E, F)。這表明GFFKR基序是將整合素α5靶向到遷移體形成位點所必需的,可能是通過影響與Rab35的關聯。
接下來,作者想直接測試Rab35和整合素α5之間可能的相互作用。使用雙色熒光相互關聯光譜(dcFCCS)捕捉Rab35與整合素α5的胞質結構域(ITGα5-cyto)之間的相互作用(圖4)。作者首先純化了GFP-Rab35-WT和GFPRab35-Q67L(組成型活性突變體)蛋白,合成了用熒光團Cy5 (Cyanine5)標記的ITGα5-cyto-WT和ITGα5-cyto-1-5A。當GFP-Rab35-WT與Cy5-ITGα5-cyto-WT或Cy5熒光團混合時,兩者均未表現出顯著的互相關信號(圖4H),說明WT-Rab35與Cy5-ITGα5-cyto-WT沒有結合。而本構活性突變體GFP-Rab35-Q67L在類似實驗條件下與Cy5-ITGα5-cytoWT表現出較強的相互關聯信號(圖4H),表明GFP-Rab35-Q67L可以結合Cy5-ITGα5-cyto-WT。相比之下,GFP-Rab35-Q67L與Cy5-ITGα5-cyto-1-5A沒有交叉相關信號(圖4H),這表明整合素α5的胞質結構域與Rab35的活性形式結合需要GFFKR基序。值得注意的是,重組GFP-Rab35-WT是從大腸桿菌中純化出來的,因此應該處于失活狀態。作為對照,作者還測試了GFP-Rab35-S22N與Cy5-ITG-α5-cyto-WT的結合,發現GFP-Rab35-S22N與Cy5-ITG-α5-cyto-WT之間沒有結合。這些數據表明,活性Rab35可以通過GFFKR基序結合到整合素α5的細胞質結構域。作者推斷,如果Rab35通過與整合素α5的GFFKR基序相互作用將整合素α5招募到遷移體的形成位點,那么將含有GFFKR基序的整合素α5衍生肽加載到細胞中應該會競爭性地抑制Rab35介導的整合素α5招募到遷移體的形成位點,從而減少遷移體的形成。事實上,作者發現用質膜滲透肽Tat-ITGα5-cyto-WT處理細胞減少了整合素對遷移體形成部位的靶向和遷移體的形成。相比之下,用GFFKR/AAAAA突變肽處理細胞未能阻斷整合素靶向或遷移體的形成(圖4I-L)。這些結果表明Rab35通過將整合素靶向到遷移體形成位點來促進遷移體的形成。
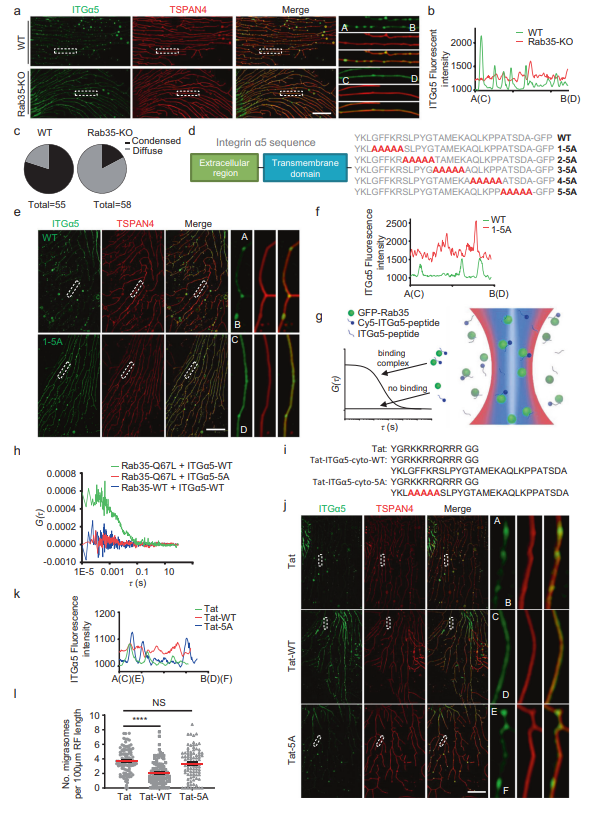
圖4 Rab35通過靶向整合素α5促進遷移體的形成
5. PI(4,5)P2-Rab35軸在生理上調控遷移體的形成,并且在進化上是保守的
最后,作者測試了PI(4,5)P2-Rab35軸是否在不同環境下調節遷移體的形成。作者首先測試了BJ細胞,這是一種從新生兒男性正常包皮皮膚上建立的成纖維細胞系。作者發現,用PIP5K1A抑制劑ISA2011B(圖5A, B)或敲低PIP5K1A(圖5C, D)處理BJ細胞,會顯著損害遷移體的形成。此外,用Tat-ITGα5-cyto-WT處理BJ細胞,可以阻斷遷移體的形成(圖5E, F)。這些觀察結果表明,PI(4,5)P2-Rab35軸對遷移體形成的調節在人類細胞中是保守的。最后,作者在體內測試了PI(4,5)P2-Rab35軸是否調控遷移體的形成。以前,作者報道斑馬魚在胚胎發育過程中形成遷移體。這里,作者發現與GFFKR/AAAAA突變肽相比(圖5K, L),用PIP5K1A抑制劑(圖5G, H)處理斑馬魚胚胎可以顯著減少遷移體的形成。此外,使用反義morpholino寡核苷酸(MO)敲除Rab35可顯著降低斑馬魚胚胎中遷移體的形成(圖5I, J)。總之,這些發現表明PI(4,5)P2-Rab35軸在一系列生理環境中調節遷移體的形成,并且在不同的脊椎動物中是保守的。
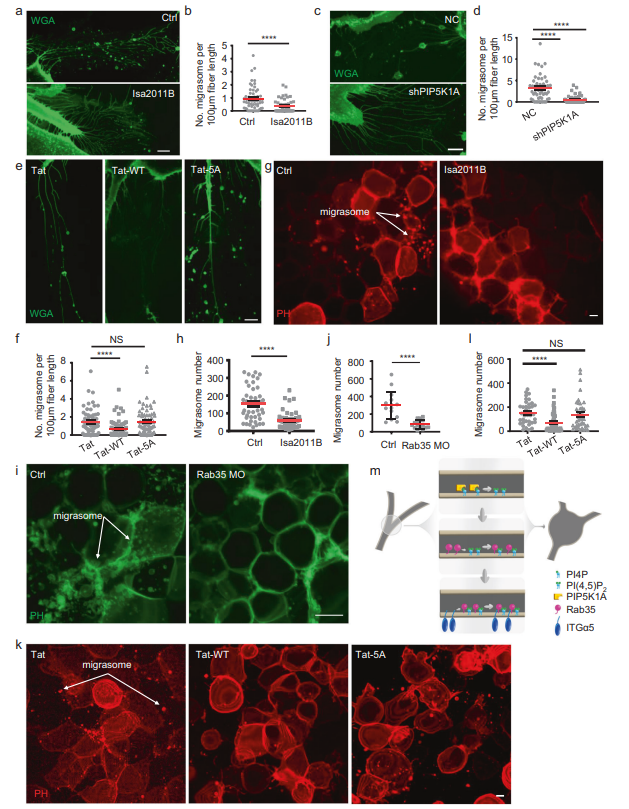
圖5 PI(4,5)P2-Rab35軸在生理上調控遷移體的形成,并且在進化上是保守的
討論
作者提出調節遷移體生物發生的信號事件的臨時模型(圖5)。PIP5K1A的募集和遷移體形成位點上PI(4,5)P2的重新合成可能是遷移體形成的觸發信號。一旦PI(4,5)P2達到濃度閾值,活性Rab35通過其多堿性簇被招募到遷移體形成部位。然后,Rab35作為適配器將整合素募集到遷移體形成位點。因此,活性Rab35和整合素之間的相互作用為遷移體的形成創造了必要的粘附點。
細胞器的生物發生通常受到信號通路的嚴格調控。在許多情況下,脂質激酶是這些信號級聯反應的核心,它將代謝、機械和其他信號結合起來,啟動特定細胞器的生物發生。這項研究結果揭示了PI(4,5)P2-Rab35軸在遷移體形成中的重要作用。遷移體可以添加到越來越多的細胞器列表中,其生物發生由磷酸肌苷信號控制。此外,作者的數據表明,遷移體的形成是一個受信號通路嚴格調控的主動生物發生過程,而不是一個膜脫落過程,在這個過程中,膜碎片被動地從遷移細胞的后緣丟失。作者發現PIP5K1A在遷移體形成之前被招募到遷移體形成的部位,這個過程中的分子機制仍不清楚,可能是由遷移體形成部位的特定脂質/蛋白質組成決定的;也有可能生物物理特性,如膜曲率,可能有助于PIP5K1A的優先招募。未來的研究需要解決這個重要的問題。PIP5K1A募集后PI(4,5)P2的快速積累,表明至少有一部分遷移體形成位點上的PI(4,5)P2是由位于這些位點的PIP5K1A重新合成的。遷移體上高度富集的PI(4,5)P2不會擴散到RFs上,這表明遷移體形成部位可能具有獨特的特性,有利于PI(4,5)P2的駐留。作者推測,從頭合成加上PI(4,5)P2的駐留可能解釋了PI(4,5)P2在遷移體形成部位的快速積累。研究證實整合素α5靶向遷移體形成位點依賴于活性Rab35。dcFCCS分析表明,整合素α5的胞質部分可以通過其GFFKR基序與活性Rab35相互作用。這些數據表明Rab35可能通過直接相互作用將整合素α5募集到遷移體形成位點。然而,Rab35與整合素α5的相互作用也有可能是間接的。除了整合素,Rab35可能還有其他效應蛋白和銜接蛋白參與遷移體的生物發生。此外,Rab35的活性如何在遷移體的生物發生中受到調節尚不清楚。為了回答這些重要的問題,未來的研究應關注遷移體生物發生的復雜調控網絡。
實驗方法
細胞培養和轉染,免疫熒光,成像和圖像分析,蛋白純化,多肽合成,ITGα5-細胞肽標記,dcFCCS測量和數據分析
參考文獻
Ding T, Ji J, Zhang W, Liu Y, Liu B, Han Y, Chen C, Yu L. The phosphatidylinositol (4,5)-bisphosphate-Rab35 axis regulates migrasome formation. Cell Res. 2023 Aug;33(8):617-627. doi: 10.1038/s41422-023-00811-5.