






在上皮性卵巢癌模型中,ODF2L作為WEE1抑制的合成致死伴侶
實驗方法:抗體和小分子抑制劑,合成致死性RNAi篩選,細胞活力測定和菌落形成試驗,細胞同步和G1/S釋放,CDK1激酶活性測定,EdU摻入試驗,蛋白過表達的質粒構建,BRET化驗
WEE1已成為上皮性卵巢癌(EOC)的一個有吸引力的靶點,但EOC細胞如何改變其對WEE1抑制的敏感性尚不清楚。本文通過細胞周期機制相關基因RNAi篩選,發現靶向精子尾部外致密纖維2樣(ODF2L)是EOC細胞中具有WEE1激酶抑制作用的合成致死伴侶。在體外和體內異種移植的EOC細胞系中,敲低ODF2L可使細胞對WEE1抑制劑AZD1775敏感。機制上,ODF2L丟失后對WEE1抑制的敏感性增加伴隨著DNA損傷的累積。ODF2L將WEE1的功能冗余激酶PKMYT1募集到CDK1-cyclin B復合物上,從而在WEE1被抑制時限制了CDK1的活性。臨床上,ODF2L的上調與患者來源的EOC細胞中CDK1活性、DNA損傷水平和對WEE1抑制的敏感性相關。此外,在EOC患者來源的異種移植模型中,ODF2L水平預測了對WEE1抑制的反應。在ID8卵巢癌同基因小鼠模型中,包裝ODF2L siRNA和AZD1775的腫瘤靶向脂質納米顆粒聯合治療可協同抑制腫瘤生長。這些數據表明,對于表達低水平ODF2L的EOC細胞,WEE1抑制是一種很有前景的精確治療策略。
技術路線:

結果:
(1) EOC細胞中ODF2L的表達與對WEE1抑制的敏感性相關
為深入了解影響EOC細胞對WEE1抑制敏感性的機制,我們使用從TRC慢病毒shRNA文庫中篩選的定制慢病毒shRNA文庫進行了細胞周期機制相關基因RNAi篩選,并針對由1,295個shRNA構建物代表的263個人類細胞周期相關基因。初步篩選涉及將A2780轉化為含有針對263個單個基因的shRNA的慢病毒池,并用亞致死劑量的AZD1775處理它們。從使用A2780細胞的初級篩選中,從263個基因中選擇前30個基因進行二級驗證篩選(圖1A)。ODF2L在初級篩選中被確定為第三個最有效的靶點,并在次級篩選中作為先導靶點出現,因為它使卵巢癌類型的癌細胞對WEE1抑制敏感(圖1B)。ODF2L的表達與卵巢癌對AZD1775的耐藥性呈正相關(圖1C)。為了進一步證實ODF2L參與了EOC對WEE1抑制的應答,我們建立了體內AZD1775耐藥模型,模擬化療耐藥的臨床發展過程(圖1D)。用A2780細胞接種裸鼠SC。在AZD1775的作用下,A2780異種移植物在最初15天內生長緩慢。然而,腫瘤的生長最終在晚期明顯增加。在終點處死小鼠進行腫瘤切除,AZD1775處理組與對照組之間腫瘤體積無顯著差異(圖1D)。相比之下,AZD1775處理后的腫瘤細胞中AZD1775 IC50比未經處理的腫瘤細胞增加了約4.5倍(圖1E),表明在體內成功誘導了對WEE1誘導的抗性。在這個體內模型中,AZD1775處理組的腫瘤細胞中ODF2L的表達顯著增加,通過RT-qPCR和免疫印跡驗證了這一點(圖1F)。在來自配對親本細胞系的4個AZD1775抗性卵巢細胞系中,ODF2L的表達一致且顯著增加(圖1G)。此外,在RNAi篩選中鑒定的30個基因中,有19個在具有WEE1抑制的特定親本細胞系中具有亞致死功能(圖1B),并且它們在配對抗性細胞系中的表達特異性增加。綜上所述,ODF2L表達與卵巢癌對WEE1抑制的敏感性呈負相關。
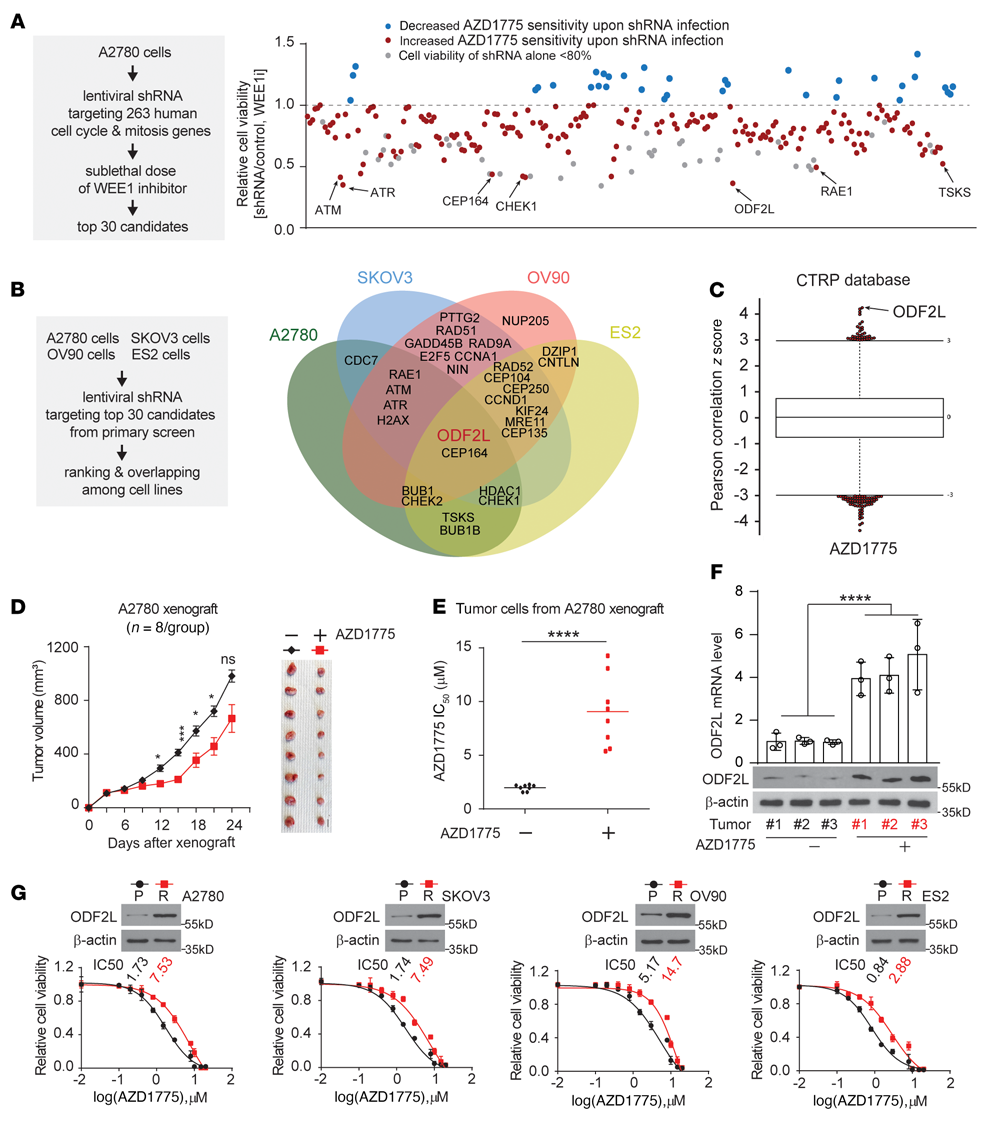
圖1:EOC細胞中ODF2L的表達與對WEE1抑制的敏感性相關
(2) ODF2L在EOC細胞中作為WEE1的合成致死伴侶
為了研究ODF2L在EOC細胞對WEE1抑制反應中的作用,我們在4個EOC細胞系中使用2個不同的shRNA克隆靶向下調ODF2L(圖2A)。雖然單獨敲除ODF2L不會顯著降低細胞活力或集落形成潛力,但在亞致死劑量AZD1775處理后,ODF2L的缺失導致細胞活力和集落形成顯著降低(圖2A, B)。此外,使用FACS, ODF2L敲除顯著加重了亞致死劑量AZD1775處理的EOC細胞的凋亡細胞死亡。進一步證實了ODF2L在EOC細胞中作為WEE1的合成致死伴侶的作用(圖2C)。此外,在異種移植小鼠模型中,還觀察到ODF2L缺失后對AZD1775處理的致敏性。在AZD1775處理的小鼠中,ODF2L敲低導致腫瘤生長和腫瘤大小顯著降低(圖2D, E)。這些數據表明,ODF2L表達顯著影響EOC細胞對WEE1抑制的反應。
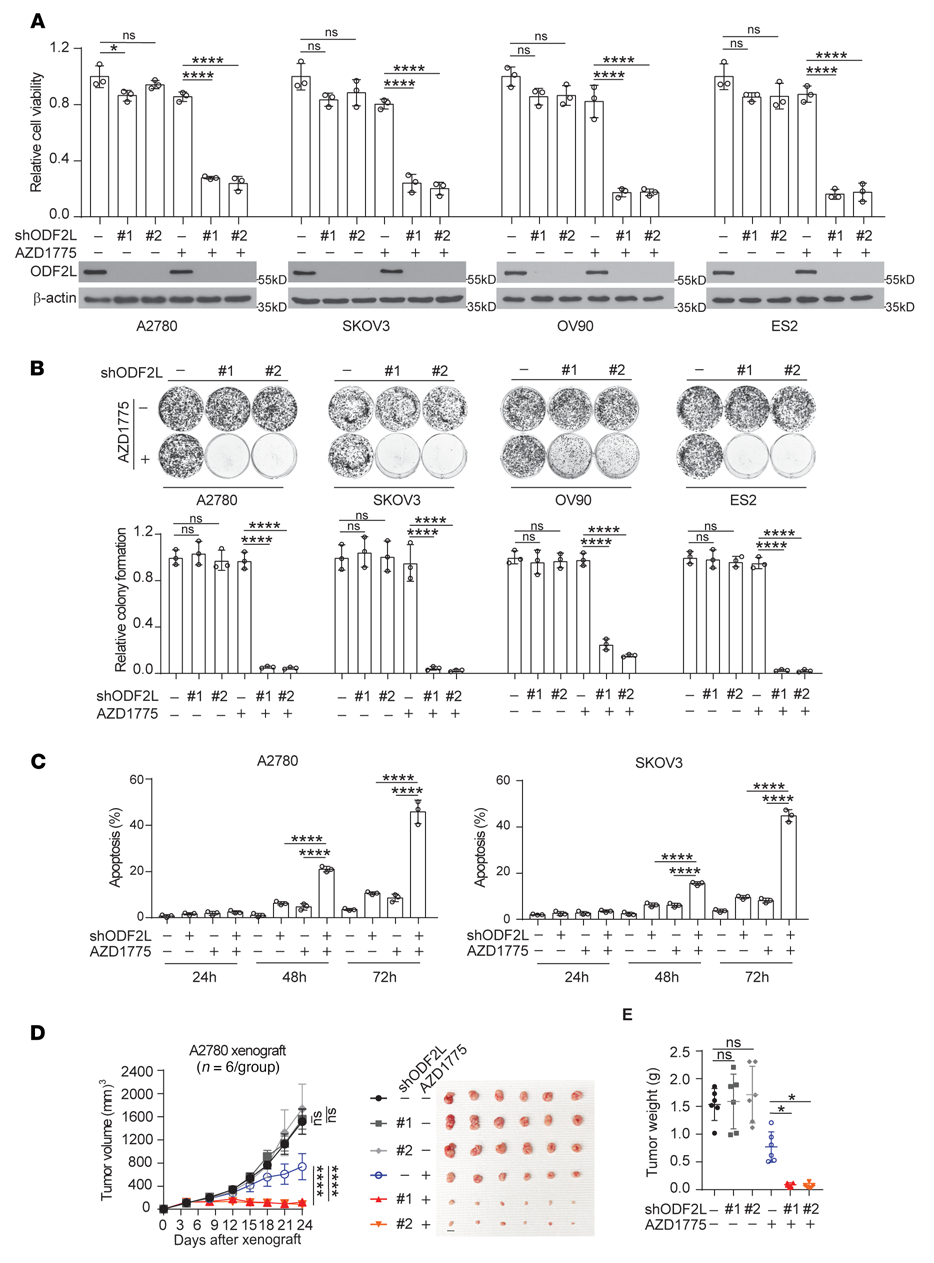
圖2:ODF2L在EOC細胞中作為WEE1的合成致死伴侶
(3) ODF2L缺失加重了WEE1抑制引起的EOC細胞DNA損傷
WEE1抑制可通過CDK1的異位激活促進有絲分裂進入并增加基因組的不穩定性。組成型活性CDK1 T14A/Y15F突變體的單等位基因表達可誘導小鼠胚胎成纖維細胞(MEFs)的復制應激和細胞死亡,并顯著增加γ-H2AX水平、染色體斷裂和DNA損傷反應(DDR)激活。為深入了解ODF2L使細胞對AZD1775敏感的機制,我們使用DSB標記物γH2AX來監測DNA損傷水平。如圖3A所示,在A2780和SKOV3細胞中,通過免疫熒光染色檢測,AZD1775處理后,ODF2L敲低顯著增加了γ-H2AX+細胞的百分比。與γ-H2AX一致,與對照細胞相比,在AZD1775處理下,ODF2L敲低的細胞中,彗星尾部的DNA數量(圖3B)和ATM、CHK2和RPA32/RPA2的磷酸化水平(圖3C)顯著增加。為了測試DNA損傷的增加是否是ODF2L敲除細胞中存在WEE1抑制的細胞周期調節的綜合缺陷的結果,我們使用基于click化學的EdU測定法結合DNA含量染色Hoechst 33342來監測細胞周期分布。如圖3D所示,AZD1775處理下,ODF2L敲低細胞中DNA含量介于1C和2C之間但EdU-(1C<DNA含量<2C/EdU-)的細胞群百分比顯著增加,這也可以通過在ODF2L敲低細胞中強制表達耐shRNA的ODF2L來挽救(補充圖未展示)。這些數據進一步支持了ODF2L缺失和WEE1抑制的結合導致非計劃進入有絲分裂,導致有絲分裂中DNA含量不完全的細胞數量顯著增加,這是一種有害的DNA損傷形式。與此結果一致的是,通過流式細胞術細胞分選后的免疫印跡檢測,DNA含量在1C和2C之間但EdU-(1C<DNA含量<2C/EdU-)的細胞群表達了大量的γ-H2AX(圖3E)。以上結果表明,ODF2L敲低可促進EOC細胞周期失調,進一步加重WEE1抑制誘導的DNA損傷。

圖3:ODF2L缺失加重了WEE1抑制引起的EOC細胞DNA損傷
(4) ODF2L抑制AZD1775誘導的CDK1活性
我們探討了ODF2L在EOC細胞中促進細胞周期失調并進一步加劇WEE1抑制誘導的DNA損傷的機制。考慮到WEE1的主要下游靶點是CDK1-cyclin B1復合物,我們首先通過監測體外CDK1活性的變化來檢測當WEE1被抑制時,ODF2L是否恢復了CDK1的失活。首先,在G1-S釋放后,用AZD1775處理ODF2L敲低或過表達的細胞和指定的對照細胞4小時。然后提取細胞裂解液并與重組CDK1底物GST-PP1Cα孵育。然后我們通過Western blotting定量pT320 GST-PP1Cα的總水平以監測CDK1活性。ODF2L敲低和WEE1抑制的結合增強了CDK1的活性(圖4A),延長了CDK1的激活狀態(補充圖未展示)。Tyr15和Thr14的磷酸化是控制CDK1活性的關鍵,這使CDK1-cyclin B1復合物在細胞接近有絲分裂前受到抑制。WEE1特異性磷酸化CDK1的Tyr15,而PKMYT1在Tyr15和Thr14上具有雙重活性。我們觀察到ODF2L的缺失影響Tyr15和Thr14的磷酸化,尤其是CDK1中Thr14磷酸化的強烈衰減,與PKMYT1的功能驚人地相似(圖4B)。為了確定ODF2L是否在CDK1 Thr14磷酸化中起重要作用,我們產生了表達CDK1 T14A-、CDK1 Y15F-或CDK1 WT-的EOC細胞,去除內源性CDK1,并監測AZD1775處理下這些EOC細胞中CDK1活性的變化。ODF2L缺失顯著增加了CDK1 WT細胞的CDK1活性,但在CDK1 T14A細胞中沒有(圖4C)。相反,當ODF2L被敲低時,在CDK1 WT和CDK1 Y15F細胞中也同樣觀察到CDK1活性的增加(圖4D),這表明ODF2L可能特異性地通過CDK1 Thr14磷酸化來抑制WEE1抑制下的CDK1活性。與WEE1抑制后的對照細胞相比,使用CDK1抑制劑Ro-3306治療可以有效地挽救ODF2L敲除細胞的集落形成潛力(圖4E),并減少凋亡細胞死亡(圖4F)。綜上所述,ODFL2可以促進CDK1 Thr14磷酸化,從而在WEE1抑制的情況下抑制EOC細胞中的CDK1活性。
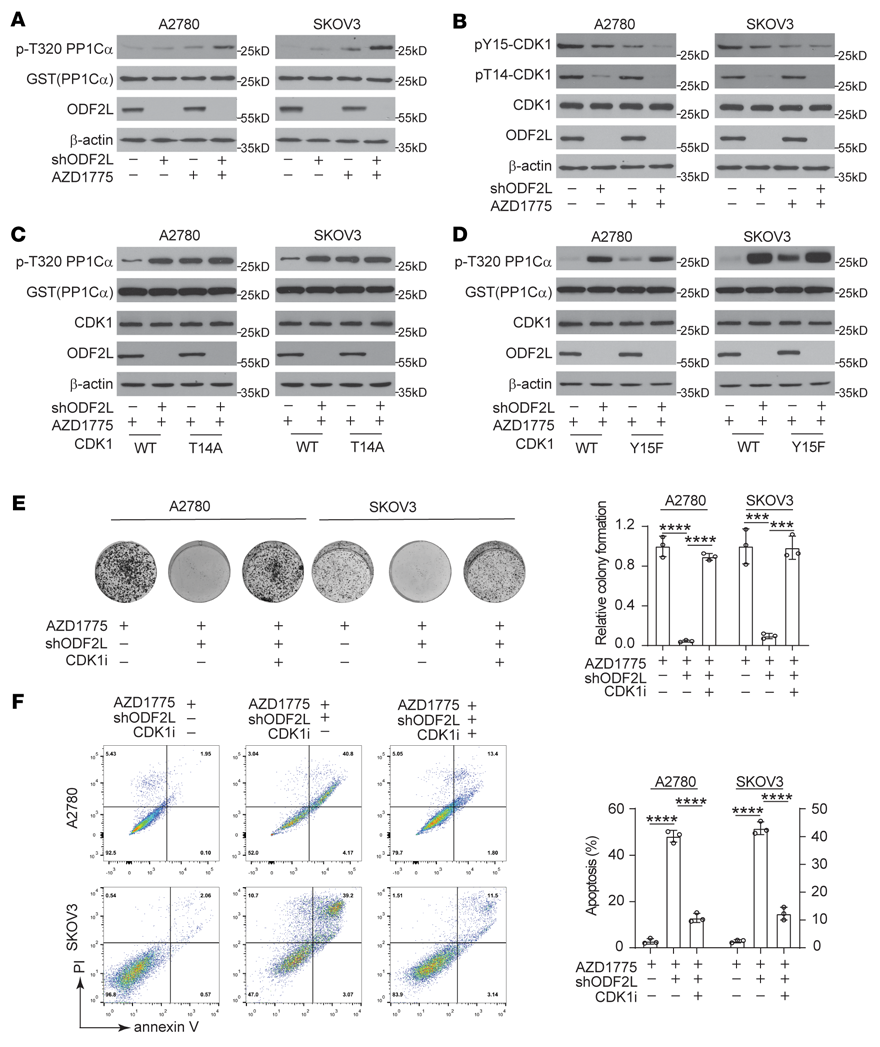
圖4:ODF2L抑制AZD1775誘導的CDK1活性
(5) ODF2L允許PKMYT1招募到CDK1復合體
我們探討了ODF2L如何在WEE1抑制的情況下促進EOC細胞中CDK1的磷酸化。與WEE1和PKMYT1水平與其底物CDK1水平之間的強相關性類似,ODF2L水平也與CDK1水平顯著相關(圖5A)。因此,ODF2L可能參與WEE1或PKMYT1復合物調控CDK1磷酸化。考慮到ODF2L在調節CDK1磷酸化Thr14 (p-Thr14)中的作用與PKMYT1重疊,我們首先測試了ODF2L是否與PKMYT1共同調節CDK1磷酸化。G1-S釋放4小時后提取EOC細胞裂解液,用于對ODF2L或PKMYT1的免疫沉淀。ODF2L和PKMYT1之間有很強的相互作用(圖5B)。ODF2L敲低在很大程度上減少了EOC細胞中CDK1免疫沉淀復合物中PKMYT1的存在(圖5C);相比之下,PKMYT1敲低對EOC細胞CDK1免疫沉淀復合物中ODF2L的水平沒有顯著影響(圖5D)。此外,在完整細胞的背景下,CDK1和PKMYT1之間的相互作用也被ODF2L的敲低所消除,這可以通過恢復ODF2L的表達來恢復,正如在共轉染了Halo-PKMYT1和Nluc-CDK1的EOC細胞中進行的生物發光共振能量轉移(BRET)實驗所示(圖5E)。在轉染了表達有Flag標記的ODF2L的n端、中程和c端結構域的質粒的EOC細胞中,通過對Flag的免疫沉淀,我們證實了ODF2L的n端和c端結構域分別直接與PKMYT1和CDK1相互作用(圖5F)。一致地,在EOC細胞中,PKMYT1的過表達或下調分別增加或降低對AZD1775的抗性,而ODF2L的下調則完全消除了這種作用(圖5G, H),這表明ODF2L在介導PKMYT1恢復G2/M檢查點方面發揮了關鍵作用。單獨敲除ODF2L比敲除PKMYT1在致敏AZD1775方面更有效,這可能是因為殘留的PKMYT1仍然可以被ODF2L募集和富集以滅活CDK1(圖5H)。綜上所述,ODF2L介導了PKMYT1與CDK1復合物的相互作用,因此當WEE1在EOC細胞中被抑制時,PKMYT1使CDK1和G2/M檢查點恢復失活。
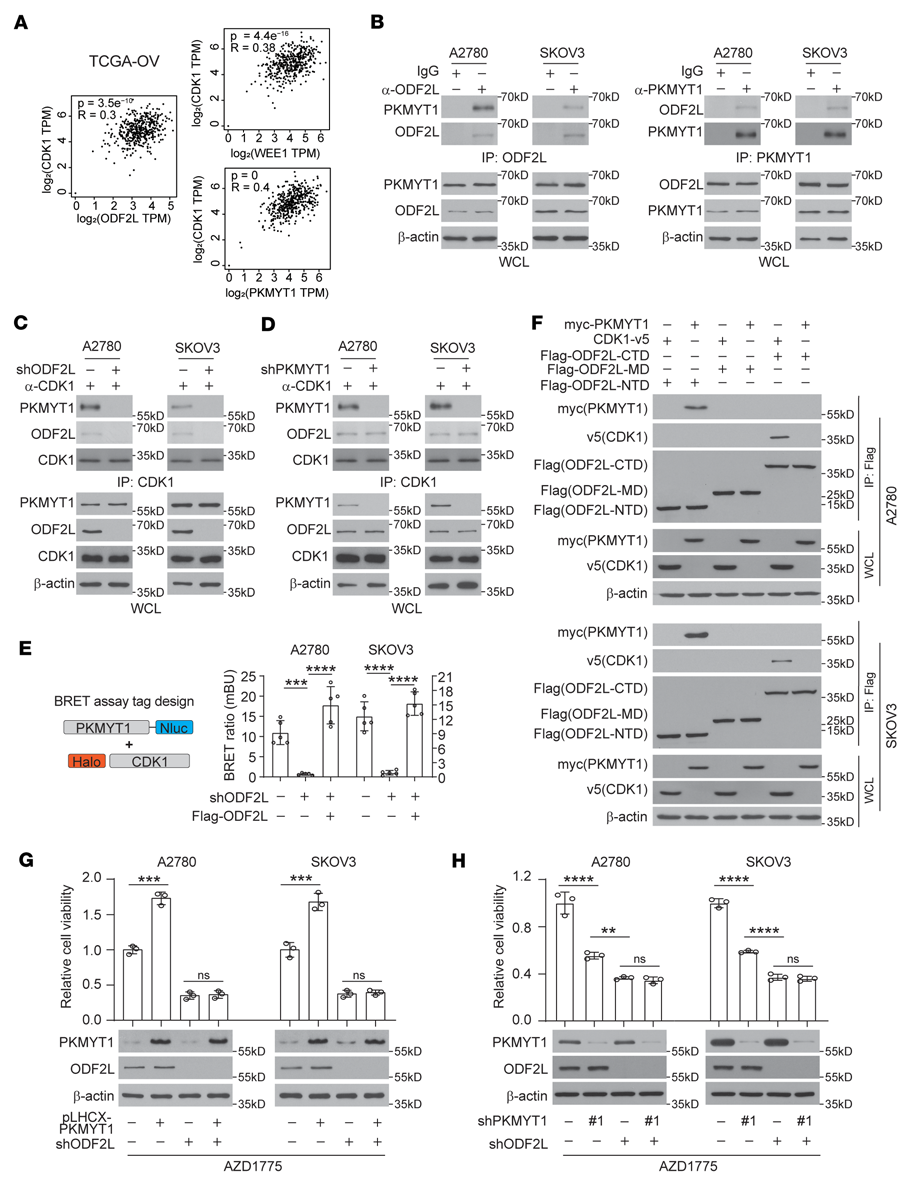
圖5:ODF2L允許PKMYT1招募到CDK1復合體
(6) ODF2L表達水平與AZD1775在EOC中的敏感性有臨床相關性
為了進一步驗證ODF2L與WEE1抑制EOC應答之間的關系,我們在一組來自57例卵巢癌患者組織樣本的AZD1775處理的原代EOC細胞中檢測了ODF2L水平、CDK1活性和細胞活力的相關性。為了檢測CDK1活性,分離的原代細胞在G1-S釋放后用AZD1775處理4小時。然后提取細胞裂解液并與重組CDK1底物GST-PP1Cα孵育。然后用Western blotting檢測細胞總裂解物中p-Thr320 GST-PP1Cα和ODF2L的總水平,并用ImageJ軟件進行定量(圖6A)。我們還監測了AZD1775處理后72小時的細胞活力(圖6B)。在AZD1775處理的原代EOC細胞中,ODF2L表達水平與CDK1活性呈強負相關(圖6C),與細胞活力呈顯著正相關(圖6D),表明ODF2L在EOC細胞對WEE1抑制的反應中起重要作用。此外,ODF2L表達水平也預測了患者源性異種移植(PDX)小鼠模型體內對AZD1775治療的不同反應。AZD1775治療在ODF2L低表達的患者癌組織中導致腫瘤生長明顯延遲,而在ODF2L高表達的患者癌組織中則沒有(圖6E)。綜上所述,這些結果有力地支持了ODF2L表達水平在臨床上與AZD1775在EOC中的敏感性相關的觀點。
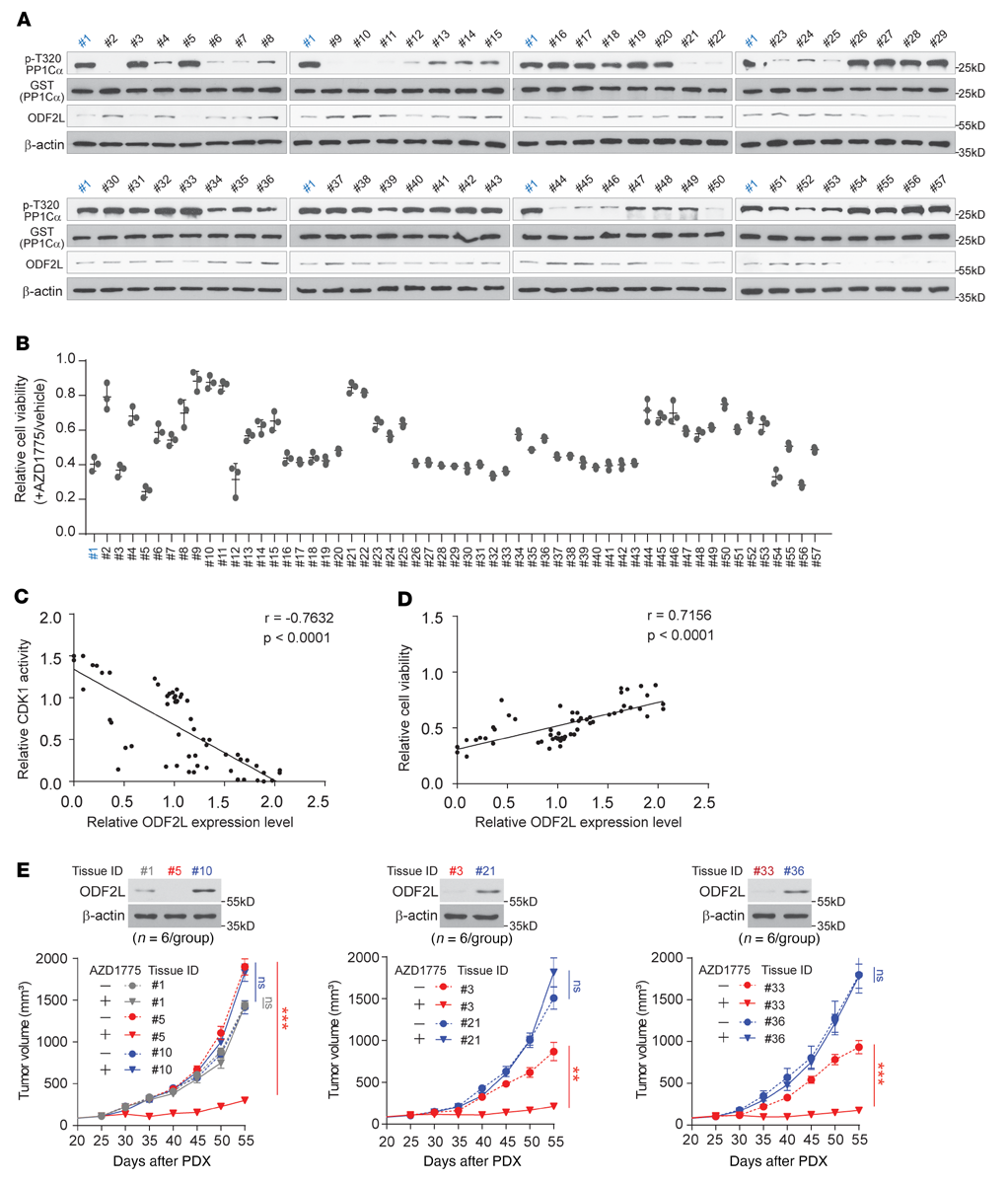
圖6:ODF2L表達水平與AZD1775在EOC中的敏感性有臨床相關性
(7) 在同基因小鼠模型中,使用RNAi治療平臺靶向ODF2L可使卵巢癌細胞對WEE1抑制劑治療增敏
ODF2L是WEE1的合成致死伙伴,ODF2L的缺失使卵巢癌對WEE1抑制劑的反應增敏,這表明ODF2L可能是卵巢癌治療的一個有希望的轉化靶點。我們利用之前建立的卵巢癌ID8Luc同基因小鼠模型,評估了靶向ODF2L聯合WEE1抑制劑的體內治療潛力。ODF2L是一種沒有市售抑制劑的支架蛋白;因此,我們使用靶向RNAi治療性脂質納米顆粒(LNP)平臺ASSET(錨定繼發性scFv靶向)在體內特異性地降低ID8癌細胞中的ODF2L。通過與識別大鼠IgG2a Fc區(ASSET)的脂質錨定單鏈抗體連接體結合,這些靶向LNPs被細胞靶向抗體包裹。由于卵巢ID8細胞特異性且高度表達EGFR,我們將負載ODF2L siRNA (siODF2L-loaded)或負載scramble siRNA (siScramble-loaded)的LNPs包被抗EGFR抗體,并將其靶向到ID8細胞(圖7A)。腹腔轉移性ID8-Luc腫瘤小鼠在腫瘤接種后7天腹腔注射與抗EGFR結合的siScramble-LNPs或siODF2L-LNPs (0.75 mg/kg),同時每天注射DMSO或AZD1775 (30 mg/kg)。采用活體生物發光成像(BLI)監測腫瘤生長情況。siODF2L-LNPs和AZD1775聯合使用可顯著抑制腫瘤生長潛力,而對照小鼠,單獨使用siODF2L-LNPs或單獨使用AZD1775均未觀察到腫瘤生長的顯著差異(圖7B, C)。一致地,總生存率顯著提高(圖7D),腹膜出血性腹水減少,ODF2L丟失和AZD1775聯合治療組腫瘤結節數減少(圖7E)。siODF2L-LNPs治療成功地降低了ODF2L在ID8腫瘤中的表達,但在主要器官,包括心臟、肝臟、脾臟、肺和肝臟(圖7F)中沒有降低ODF2L的表達(圖7F),導致腫瘤細胞中CDK1的大量激活(圖7F)和DNA損傷加劇(圖7G)。這些數據為ODF2L與WEE1抑制劑在卵巢癌治療中作為有效的合成致死靶標伴侶提供了原理證明。
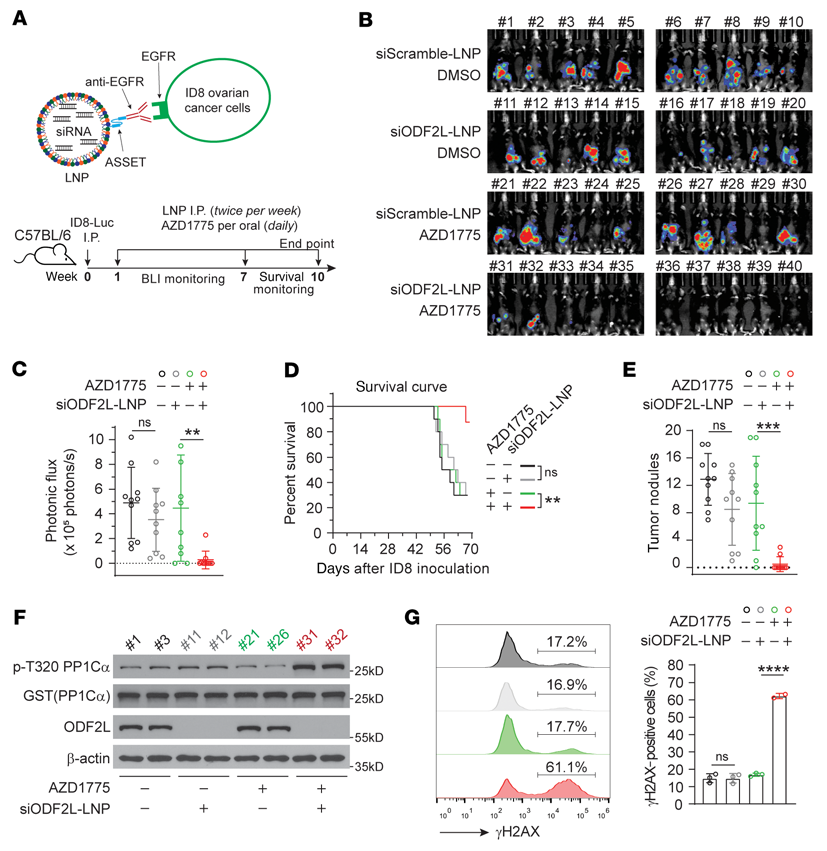
圖7:在同基因小鼠模型中,使用RNAi治療平臺靶向ODF2L使卵巢癌細胞對WEE1抑制劑治療增敏
結論:ODF2L是WEE1的合成致死伴侶,在恢復人類EOC細胞中AZD1775抗性G2/M細胞周期檢查點信號通路中起關鍵作用。臨床上,ODF2L可作為一種預測標志物,并作為一種有希望的治療靶點,聯合WEE1抑制劑治療EOC患者。
參考文獻:Li, J., Lu, J., Xu, M., Yang, S., Yu, T., Zheng, C., Huang, X., Pan, Y., Chen, Y., Long, J., Zhang, C., Huang, H., Dai, Q., Li, B., Wang, W., Yao, S., & Pan, C. (2023). ODF2L acts as a synthetic lethal partner with WEE1 inhibition in epithelial ovarian cancer models. The Journal of clinical investigation, 133(2), e161544. https://doi.org/10.1172/JCI161544.